Concept
森林再生テクニカルノート(TPPs)は、途上国の劣化が進んだ森林や開発後に放棄され荒廃した土地等において、効果的な森林の再生に大きく貢献する技術集です。
by ![]()
Technical Note for Tree Planting Practices 森林再生テクニカルノート
海外植林活動のための技術データベース
樹幹穿孔性蛾類(ボクトウガ科・コウモリガ科)
樹木の幹に穿孔する蛾類(鱗翅目)のうち特に大きな被害を及ぼす樹幹穿孔性蛾類として、チークビーホールボーラー、ゴマフボクトウ属(Zeuzera)、コウモリガ属(Endoclita)を中心に述べる。
チークビーホールボーラー



分類・分布
チークビーホールボーラー(Xyleutes ceramicus)は、チーク(Tectona grandis)の穿孔性害虫として古くから知られている。学名をDuomitus ceramicusとすることもある。ビーホール(bee hole)という呼称は、本種の幼虫の坑道がハチの穿孔によるものとされたことに由来する。ミャンマー、タイ、マレーシア、インドネシア、ニューギニア、ソロモン諸島などに分布する。本種の被害はミャンマーとタイで多く、最近チーク造林が盛んになったマレーシア(半島部およびサバ州)でも被害が目立つようになっている。しかしインドネシアのジャワ島では被害は比較的少ない。シッキムなどヒマラヤ山脈東部以外のインドの大部分には本種は分布していない。
形態・生態
成虫は横長の翅型でサイズはメスがオスより大きくまた個体差があり、翅開長はオス40〜80 mm、メス40〜160 mm。淡褐色、濃褐色、黒色の細い横縞状の斑紋が多数ある。メス成虫は1週間ほど生存し、主に粗皮下に100〜数千卵の卵塊を産み付ける。1メスあたり総産卵数は平均12,483卵、個体によっては50,000卵という報告もある。卵は長径0.75 mm。孵化した幼虫は絹糸を吐いて風に乗って分散したり樹幹上を歩き回ったりして樹皮に食入し1〜2齢期を樹皮内で過ごす。その後辺材表面を摂食し、さらに場所を変えつつ材内へ穿孔を繰り返した後、定着場所を定め、心材に向かって斜め上方に進み、心材で垂直上方に進んで住処となる坑道を形成し、入り口付近の辺材に形成されるカルスを食べて成長する。幼虫には体節ごとに白色と桃色が交互する縞模様があり老熟幼虫の体長は 60〜70 mm前後に達する。坑道内で頭を下に向けて蛹化し、蛹化に先立って坑道の入り口を絹糸の塊で閉じる。タイでは雨季(植樹の成長期)に幼虫期が同調し、成虫は乾季の終わり3月に出現、産卵し、幼虫は乾季の始まる11月ごろに蛹化する。ジャワ西部やボルネオなどの降雨林地域では季節性が見られなくなる。
加害樹種
チーク(Tectona grandis)のほか、キダチヨウラク(別名:ヤマネ、グメリナ)(Gmelina arborea)、グローリーツリー(Clerodendrum infortunatum、Callicarpa arborea、Vitex parviflora)等のシソ科(以前クマツヅラ科とされた)に加害する。このほか、ノウゼンカズラ科のカエンボク(Spathodea campanulata)、ミソハギ科のDuabanga grandifloraおよびD. Sonneratoides等にも加害の記録がある。
被害
後期齢の幼虫が材内深く穿孔することにより、材価が損なわれる。早いものでは2年生、胸高直径10cm未満から被害が発生する。幼虫の坑道は25 cm程度に達する。幼虫がいる坑道の入り口からはフラス(糞)が排出されるが、成虫が羽化した後はカルスの発達によって入り口が塞がれるので外からは被害がわかりにくい。タイのチークでの調査では被害部位は食樹の成長とともに高い位置に移る傾向があり、6年生で0〜1 m、9年生で2〜14 m、14年生で3〜15m、30年生で8〜11mという報告がある。また、タイでは成長の良い林分で被害が多く、年間降雨量が1750〜2750 mmのところで被害が多いという。また、ジャワでは雑草が繁茂している林分で被害が多いという。
防除
- 化学薬剤
- 生物的防除
- フェロモン
- 抵抗性
- その他
穿孔性であるため化学薬剤散布の効果は低い。薬剤を坑道に注入すれば確実に幼虫を殺せるが、対象本数が少ない場合に限定される方法である。
ボーベリア菌(Beauveria bassiana)および市販のBT剤(Bacillus thuringiensis Berliner)が有効であったという試験結果がある。捕食者としてキツツキ類、アリ類が有力であると言われ、火災や火入れはアリ類の生息を困難にするので避けるべきであるとの指摘がある。
性フェロモン候補物質が特定されている。今のところ実用段階にはないが、将来はモニタリング、誘殺、更新撹乱への利用が期待される。
タイのチークでは被害の少ない系統があり、抵抗性育種の可能性がある。被害が少ない理由は、樹皮が薄く乾いているので若齢幼虫の生育に不適なためだという。
粗皮を掻き落とすことで若齢幼虫が排除でき、坑道の入り口をナイロンネットで塞ぐことで羽化した成虫の脱出を阻止することもできる。労力を投入できるなら生活環が一定している季節林地帯で適期を狙って実施することも考えられる。
補足
Dervishiya cadambae (Moore)はインドでチークを加害するボクトウガ科の1種で、Alcterogystia cadambae、またはCossus cadambaeとされたこともある。チークの大径木を加害し、それが原因で枯死に至ることもある。チーク以外にはNeolamarckia cadambaも加害する。
ゴマフボクトウ属

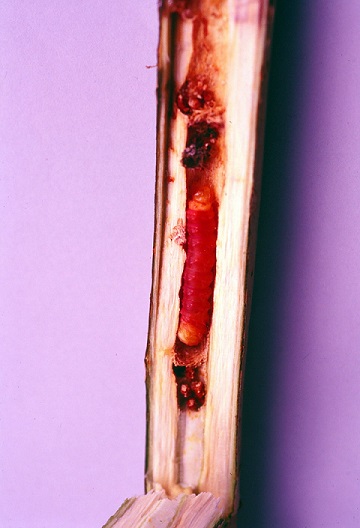
分類・分布
ゴマフボクトウ属(Zeuzera)はユーラシア・北アフリカ・オセアニアの熱帯・亜熱帯・暖温帯に10数種が知られているが、これらをいくつかの属に細分する意見もある。多食性で多くの科にわたる樹種の幹や太枝を穿孔加害する。このため果樹・庭園樹・造林木などさまざまな有用樹木の害虫として知られている。熱帯地域で樹木害虫として記録されているのはコーヒーゴマフボクトウ(Zeuzera coffeae Nietner)で、インド、東南アジア、オセアニア、台湾、中国南部に分布するが、アジアの熱帯には互いによく似た本属の種が多いので、本種のみ被害記録が特に多いのは同定が正しくないためかもしれない。中東から地中海地域にかけてはZ. pyrina L.が果樹園の重要害虫であり、日本ではゴマフボクトウ(Z. multistrigata Moore、またはZ. leuconotum Butler)とされていたこともある)が、茶畑や緑化樹・庭園樹の害虫としてよく問題になっている。これらの種では生態や防除法の研究が進んでいる。
形態・生態
本属も成虫は横長の翅型であり、白い地色に黒い点状の斑紋が散在する。大きさは種によって多少異なり、メスはオスより大型。翅開長は大型種ゴマフボクトウの大型のメスで70 mm程度、小型種のコ―ヒーゴマフボクトウのオスで30 mm程度。メス成虫は枯れ枝や樹皮の裂け目などに卵塊を産み、日本産のゴマフボクトウでは1卵塊あたり平均101卵、1メスあたり総産卵数は318〜1800卵という報告がある。台湾のコーヒーゴマフボクトウでは総産卵数190〜1134卵という報告がある。孵化した幼虫は絹糸を吐いて風に乗って分散する(バルーニング)。初め新梢に食入し、その後木化した茎や幹に入り、数回食入部位を変えながら成長する。成長した幼虫は赤褐色で、ゴマフボクトウでは 35-50 mm、コーヒーゴマフボクトウでは約30-40 mm。ゴマフボクトウでは最後に根元近くに入り樹幹や根部を穿孔するというがコーヒーゴマフボクトウやZ. indicaでは老熟幼虫でも地際より高い位置にいて根に入り込まないものも多い。この段階では食入孔から排出した褐色粒状の糞が地際に溜まるので良く目立つ。坑道の中で蛹化する。
加害樹種
本属は一般に多食性であり、多くの広葉樹を加害する。アジア熱帯のコーヒーゴマフボクトウは造林樹種ではユーカリ類(Eucalyptus spp.)、マホガニー類(Swietenia、Khaya、Toona、Cedrela)、チーク、キダチヨウラク、モルッカネム(Falcataria moluccana)、 Acacia mangium等に被害記録があり、コーヒー、カカオ、ワタ、グアバ等の害虫としても知られ、その他非常に多くの木本植物を加害した記録がある。このほかZ. confertaがSonneratia apetalaを加害した例、Z. indica Herrich-Scafferがマホガニーを加害した例、未同定種のZeuzera sp.がユーカリを加害した例などがある。中東からヨーロッパにかけて生息するZ. pyrinaは果樹(リンゴ、ナシ、モモ、クルミ等)、オリーブの害虫として知られている。日本のゴマフボクトウ(Z. multistrigata)は茶での大きな被害が報告されているほか、庭園樹、緑化樹の害虫としても知られ、ツバキ、サルスベリ、ブナ科、バラ科、ヤナギ科などを加害する。
被害
造林地では2〜3年生程度の幼齢木に被害が多く、幹の中に坑道を掘って進むため被害部より先が枯れるか木全体が枯れる。新梢が加害されたのち複数の新梢が再生して樹形が悪化する被害も発生する。果樹園やコーヒープランテーション等での被害は多いが造林地では通常散発的な被害にとどまる。しかし、バングラデシュのマングローブ植林ではZ. confertaによるSonneratia apetalaの三分の一以上に被害が出ている。マレーシアのサバ州とインドネシアのスマトラでコ―ヒーゴマフボクトウにより、またインドネシアの中部カリマンタンおよび東カリマンタンでは未同定のZeuzera sp.によりユーカリ類造林地で大きな被害が発生した事例があり、ユーカリは被害を受けやすいのかもしれない。
防除
- 化学薬剤
- 生物的防除
- フェロモン
穿孔性であるため温帯地域では薬剤の散布は若齢幼虫が新梢に食入する時期に合わせて行わないと効果がないと言われるが、熱帯の通年発生している地域や発生が不斉一な地域では、散布適期を特定することは困難である。被害本数が比較的少ない場合には薬剤を食入孔から注入する方法は材価が問題にならない果樹などでは推奨できるが、造林地では穿孔被害があればすでに材価が失われているので意味はない。造林地では幼齢木が被害を受ける場合が多いので、被害率が高くなければ被害木を除去し補植する方が良い。
中東でZ. pyrinaを対象とした研究がある。イランでは昆虫病原性線虫であるSteinernema carpocapsaeが有望であるとの試験結果がある。トルコではボーベリア菌(Beauveria bassiana)が有効であったという試験結果がある。エジプトでは核多角体ウイルスの利用なども検討されている。
Zeuzera pyrinaの性フェロモン成分は明らかにされており、合成フェロモン剤がすでに製品化されて、中東やヨーロッパの果樹園やオリーブ農園でトラップを用いたモニタリングや誘殺に活用されている。またフェロモンディスペンサーを使った交信撹乱も実施され、効果をあげている。インドネシアの中部カリマンタンのユーカリ造林地でZ. pyrinaのフェロモンルアーを用いた粘着式トラップを設置したところ、多数のZeuzera sp.成虫が捕獲されたという報告がある。
なお、中東・ヨーロッパでは果樹等を中心にZeuzera属に対して実施された防除の成果報告が多いが、生物的防除とフェロモンによる誘殺や更新撹乱を併用した総合防除を行うべきだとする意見も多い。熱帯の造林地でも本属に限らず樹幹穿孔性蛾類にこのような方法を援用する余地は多いと思われる。
コウモリガ類コウモリガ科




分類・分布・加害樹種
コウモリガ類(Endoclita spp.)コウモリガ科(Hepialidae)は60種ほどが南アジア、東南アジア、東アジアに分布する。情報の多い種では加害樹種が多く、広葉樹だけでなく針葉樹を加害することもある。おそらくほとんどの種が多食性であろうと予想される。また、類似種が多く、誤同定に基づく分布記録や被害記録もあると予想されるが、造林樹種の被害が知られている種は以下の通りである。
- Endoclita aroura Tindale
- Endoclita auratus Hampson
- Endoclita gmelina Tindale
- Endoclita hosei Tindale
- Endoclita malabaricus (Moore)
- Endoclita signifier (Walker)
マレーシアのサバでチーク、キダチヨウラクの被害が報告されている。
ミャンマーで、スギ(Cryptomeria japonica)、ユーカリの被害が報告されている。
半島マレーシアでチーク、キダチヨウラクの被害が報告されている。
マレーシアのサバでユーカリ(Eucalyptus deglupta)、キダチヨウラクの被害が報告されている。
インドでチーク、キダチヨウラク、ユーカリ、モルッカネムの被害が報告されている。
インド、ミャンマー、タイでチーク、中国南部でユーカリの被害が報告されている。なお日本のキマダラコウモリ(E. sinensis (Moore))はかつて本種とされていたため海外の文献では本種が日本に生息するとされていることがある。
形態・生態
大型のコウモリガ科で、成虫はやや幅広の横長の翅を持ち、翅開長は50〜100 mm程度。前翅に褐色、灰褐色、緑褐色、黄褐色などの濃淡斑紋を持つものが多いが前後翅ともほぼ一様に暗褐色の種もある。メスは大量の卵を飛びながら地上に撒くが、歩きながら産卵することもある。孵化した幼虫は若齢期を地表で過ごし、落ちている枯れ枝等に食入する種や、地面に接している草本の茎を摂食する種がある。落枝に生じた菌類を食べているものもあると言われる。中齢期以降の幼虫は樹木の幹や草本の茎に穿孔するものが多いが、地下部の根に入る種もある。幹では坑道を下向きに掘り進み、入り口は大量の糞と木屑を絹糸で綴って塞ぐが入り口から水平方向にも環状に食害する。幼虫はやや細長い体型で体節背面にキチン化した板(硬皮板)を持つ。老熟幼虫は70〜80 mmに達する。蛹化は坑道内で行う。
防除
若齢幼虫は地表付近で生活するため地表への薬剤散布は効果があることが、日本のコウモリガ類で確かめられている。中齢期以降の樹幹に穿孔している幼虫に対しては化学薬剤の注入は有効であるが、大規模造林地では被害木の特定に労力がかかる。コウモリガ類は一般にあまり高密度にはならないので、効率の悪い防除を行うより被害を許容すべきだという考えもある。コウモリガ類は独特の交尾システムを持ち、Endoclita属でも日本のコウモリガ(E. excrescens Butler)ではオスが群飛しながらメスを誘引することが知られており、雄性フェロモンも解明されている。防除にどのように利用できるかは今後の課題である。
補足
Sahyadrrassus malabaricusはインドに生息するコウモリガ科の1種で、多食性であるが造林木ではチークの幼齢木を加害する。 Endoclita属の種に似ているが、大径木は好まず、中齢以降の幼虫が幼齢木の髄に穿孔する。若齢期の生態は不明であるが地表部で生活しているものと予想されている。
参考文献
- Amani Mannakkara (2006) Red stem borer, Zeuzera coffeae (Lepidoptera: Cossidae): emerging threats to forest plantation in Sri Lanka. The Sri Lanka Forester 29: 61-67.
- Ashtari, M., J. Karimi, M.R. Rezapanah, M. Hassani-Kakhi (2011) Biocontrol of leopard moth, Zeuzera pyrina L. (Lep. : Cossidae) using entomopathogenic nematodes in Iran. IOBC/wprs Bulletin 66: 333-335.
- Atkinson, D. J. (1931) Insect damages to the timber of teak (Tectona grandis). Burma Forest Bulletin 26: 3-4.
- Atkinson, D. J. (1936) A survey of the damage to teak timber by the beehole borer (Xyleutes ceramica Wlk.) throughout the main teak bearing forests of Burma (Lepidoptera, Cossidae). Indian Forest Records 2(1): 1–98.
- Beeson, C. F. C. (1921) The beehole borer of teak-A preliminary note on ecology and economic status of Duomitus ceramica Wlk. in Burma. Indian Forest Records 8(3): 229–333.
- Chaiglom, D (1966) Teak beehole borer and the control research in Thailand. Natural History Bulletin of the Siam Society 21: 156-160, 4 plates.
- Dhanarajan, G. (1976) Some observations on the teak collar ring borer – Endoclita gmelina (Lepidoptera: Hepialidae) in north western Malaysia. Malaysian Forester 39: 214-223.
- Eungwijarnpanya, S., K. Nakamuta, T. Ikeda & C. Hutacharern (1990) Bionomics of the teak beehole borer, Xyleutes ceramicus, in northern Thailand: mating behavior. Thai Journal of Forestry 9: 196-202.
- Eungwijarnpanya, S. et al. (1994) Bionomics of teak beehole borer (Xyleutes ceramicus) in northern Thailand: Emergence and response to light trap of adult moth. Thai Journal of Agricultural Science 27: 1–8.
- 後藤忠男 (1994) タイ国における熱帯有用樹の穿孔性害虫.熱帯林業 30: 30-37.
- Gotoh, T., J.R. Kotulai & K. Matsumoto (2003) Stem Borers of Teak and Yemane in Sabah, Malaysia, with Analysis of Attacks by the Teak Beehole Borer (Xyleutes ceramica Wlk.). JARQ 37 (4): 253 – 261.
- Gotoh, T. et al. (2007) Emergence, oviposition and larval behaviors in the teak beehole borer (Xyleutes ceramica Wlk.) in northern Thailand (Lepidoptera: Cossidae). JARQ 41 (4): 307 – 314.
- Grehan, J.R. and V.A. Ismavel (2017) Forest ghost moth fauna of northeastern India (Lepidoptera: Hepialidae: Endoclita, Palpifer and Hepialiscus). Journal of Threatened Taxa 9: 9940-9955.
- Ismail, II, N.A. Abu-Zeid, F.F. Abdallah (1992) Population dynamics of the leopard moth, Zeuzera pyrini L. and its control on olive trees in Egypt. Journal of Plant Diseases and Protection 99: 519-524.
- Kan, E., T. Hidaka, T. Sato and H. Kitajima (2002) Copulation and male calling in the swift moth, Endoclitha excrescens (Butler) (Lepidoptera: Hepialidae). Applied Entomology and Zoology 37: 163-169.
- 前藤薫 (1988) タイ国におけるチークの穿孔性害虫ビーホールボーラー.森林防疫 37(4): 13-17.
- Maeto, K. & S. Eungwijarnpanya (1994) Density and spatial distribution pattern of current emeregence holes of the teak beehole borer, Xyleutes ceramicus, in northern Thailand. Thai Journal of Agricultural Science 27: 83-95.
- Marukawa, K. and K. Mori (2002) Pheromone synthesis CCXVIII. Synthesis of (1R, 3S, 5S)-1,3,8-Trimethyl-2,9-dioxabicyclo [3.3.1] non-7-ene, the male pheromone of a Hepialid Moth, Endoclita excrescens, and its Enantiomer. European Journal of Organic Chemistry 23: 3974-39768.
- Mathew G. (1990) Biology and Ecology of the teak trunk borer Cossus cadambae Moore and its possible control. Kerala Forest Research Institute Research Report 68: 1-41.
- Nair, K.S.S. (1982) Seasonal incidence, host range and control of the teak sapling borer, Sahyadrrassus malabaricus. Kerala Forest Research Institute Research Report 16: 1-36.
- Öztop, A., Keçeci, M., Tekşam, İ., & Ünlü, A. (2016). Investigations on control of Leopard Moth Borer (Zeuzera pyrina L. (Lepidoptera: Cossidae) in pomegranate orchards of Antalya province. Horticultural Studies 33: 57-68. http://doi.org/10.16882/derim.2016.74352
- Rohani, M. and M.A. Samih (2012) The efficiency of pheromone traps in attracting and capturing Zeuzera pyrina L. (Lepidoptera: Cossidae) in walnut orchards. International Journal of AgriScience 2: 583-587.
- Sajap, S. A. (1989) Incidence of attack by beehole borer, Xyleutes ceramica Wlk., in Gmelina arborea Roxb. plantation in Peninsular Malaysia. In Regional symposium on recent development in tree plantations of humid/subhumid tropics of Asia, Universiti Pertanian Malaysia, 1–6.
- Sarto i Monteys, V. (2001) Control of leopard moth, Zeuzera pyrina L., in apple orchards in NE Spain: mating disruption technique. IOBC/wprs Bulletin 24: 173-178.
- Sein, U K. (1963) The bee-hole borer of teak (Xyleutes ceramica Wlk.). The Burmese Forester 13 (2): 32-39.
- Suheri M., N.F. Haneda, Y.H. Jung, S. Sukeno, H.K. Moon (2020) Effectiveness of pheromone traps for monitoring Zeuzera sp. (Lepidoptera: Cossidae) population on Eucalyptus pellita plantation. IOP Conference Series: Earth and Environment Science 468 012016 doi: 10.1088/1755-1315/468/1/012016
- Yasuda, T. and K. Abe (1986) Endoclita hosei (Lepidoptera: Hepialidae) attacking Eucalyptus in Sabah, with description of the immature and imaginal stages. Applied Entomology and Zoology 21: 417-423.