Concept
森林再生テクニカルノート(TPPs)は、途上国の劣化が進んだ森林や開発後に放棄され荒廃した土地等において、効果的な森林の再生に大きく貢献する技術集です。
by ![]()
Technical Note for Tree Planting Practices 森林再生テクニカルノート
海外植林活動のための技術データベース
大発生を繰り返す食葉性蝶・蛾類(鱗翅目)
幼虫が樹木の葉を食害する蝶・蛾類は非常に多いが、大発生して林内の木が葉を全て失うほどの食葉害を起こす種は多くなく、それが高頻度で起こる種は比較的限定される。さらに、熱帯林は温帯林や亜寒帯林に比べて昆虫の大発生が少ないとも言われている。また食葉性鱗翅目の大発生は見た目にも強い印象を与え、林業や森林の健全性に及ぼす影響も甚大であろうとの危惧・懸念を生じがちな現象であるが、実質的な被害はないと判断できる場合もある。熱帯で大発生が比較的頻繁に起こる食葉性蝶・蛾類のうち特に被害が大きい2種について述べ、大発生の頻度は高いが比較的影響の少ない種についても補足的に述べる。
1.チークディフォリエーター(Hyblaea puera Cramer)
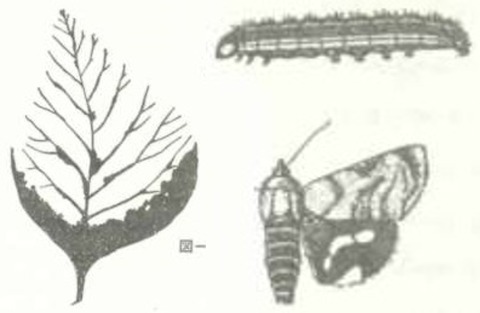
分類・分布
チーク(Tectona grandis)の食葉性害虫である、セセリモドキガ科(Hyblaeidae)のチークディフォリエーター(和名:キオビセセリモドキ、英名:teak defoliator)(Hyblaea puera)は古くから知られている。インド、東南アジア、ニューギニア、オーストラリア、オセアニアの島嶼、アフリカ、中南米など世界の熱帯・亜熱帯のほぼ全域に分布する。これらの全てが自然分布か、また全ての地域の個体群が同一種か否かについては論議もある。本種は移動力の強い種で、日本でも南の地域から飛来したと考えられる個体が稀に見られ、キオビセセリモドキという和名がある。アメリカ合衆国の南部でも発見されている。
形態・生態
成虫の翅開長は30〜40 mm、前翅は灰褐色の地色に濃褐色の斑紋があり、後翅は濃褐色の地色で中央に黄色い帯状斑紋、外縁にも黄色い斑紋がある。メス成虫は1週間ほど生存し、寄主植物の葉に1卵ずつ卵を産み付ける。1メスあたり総産卵数は約500卵、最高で1,000卵という報告もある。孵化した幼虫は葉縁の一部に切れ込みを入れて折り絹糸で閉じ合わせてその中に隠れて摂食する。幼虫は灰緑色で背中が橙黄色、あるいは全体に黒っぽい。黒化した個体は個体群密度が上昇した時に多くなる。老熟幼虫の体長は 35〜745mm前後に達する。絹糸を吐きながら降下して地上のリター中に繭を紡いで蛹化するが、降雨が多い条件やマングローブ林では食樹や下層の植物の葉を綴って繭を紡ぎ蛹化する。発育期間は温度に依存し、南インドでの野外飼育では、幼虫期間10〜12日、蛹期間6〜7日、産卵から成虫羽化まで18〜19日という報告があり、北インドでは最長36日という報告がある。年間10数世代を繰り返していると見られる。冬のある地域で越冬しているかどうかは不明であるが、明らかな冬季および夏季休眠は知られていない。
加害樹種
多くの寄主植物が記録されているが、チークのほか、キダチヨウラク(別名:ヤマネ、グメリナ)(Gmelina arborea)、グローリーツリー(Clerodendrum infortunatum、Callicarpa arborea、Vitex parviflora等)のシソ科(以前はクマツヅラ科とされていた)種への加害が多い。さらにキツネノマゴ科のヒルギダマシ属(Avicennia)、ノウゼンカズラ科のTabebuia pentaphyllaなどのシソ目植物への加害が多い。このほかクルミ科、モクセイ科、ヒルギ科なども加害したという記録がある。
被害
チーク造林地での大発生はインド、バングラデシュ、スリランカ、ミャンマー、タイ、インドネシアで知られており、コスタリカ、ブラジルなどでも大発生が起きている。また、タイ、コスタリカ、ブラジルではマングローブ林でも大発生が起きている。しかし、中南米での大発生は比較的最近の現象であり、またアフリカでは大発生の記録はない。
インド南部ケララ州のチーク造林地で大発生の経過と被害実態の詳細な調査結果の報告がある。それによると雨季に入り葉が伸び始めるとチークへの成虫の飛来・産卵が始まり、大面積の大発生が突然起こるが、その約1ヶ月前に0.5〜1.5 ha程度の小規模で先行的な大発生があると言う。さらに1〜2回の発生ピークを繰り返す年もあるが、やがていなくなる。被害木が枯れることはあまりないが、新葉展開期の食葉害による材積成長の損失は大きく、44%にも上るという調査結果がある。
本種は休眠せず、乾季にはチーク以外の植物に依存していると考えられる。また、成虫は移動性があり、集団で一定方向に向かって飛ぶという報告がある。また、大発生時に羽化した成虫はその場で産卵せず、移動してから産卵するという報告もある。近距離の林分間移動のほか、長距離移動の可能性もあり、北インドではより低緯度の地域からの移動個体の飛来により発生しているのではないかと考えられている。降雨の開始に続く新葉展開期に始まる大発生は、ミャンマー、タイ、バングラデシュ、スリランカ、インドネシアでもほぼ毎年起きている。
防除
1) 化学薬剤
化学薬剤は大面積の空中散布を含め、接触剤(現在使用が規制されているものも含む)やazadirachtinなどの散布実施例が多く、概ね効果は高いと報告されている。しかし、移動性の高い本種に一時的な効果しか期待できない薬剤散布を行うのは非効率であることと、大規模な使用には安全性への懸念があることから化学的防除には否定的な意見もある。
2) 生物的防除
本種に特異的な核多核体ウイルスHpNPV (Hyblaea puera nuclear polyhedrosus virus)が発見され、野外試験でも有効性が検証されている。ただし、降雨があると効果が落ちるという報告もあり、本種の発生が雨季に同調していることから適用法にはさらに改善が必要かもしれない。また、BT剤(Bacillus thuringiensis Berliner)が有効であったという試験結果もある。
性フェロモンは特定されていないが、仮に利用可能になっても本種の移動性が高いことから用途はモニタリング等に限定されるであろう。初期の小規模発生の段階で化学薬剤か上記生物農薬の適用による防除を行なってその後の大発生を抑制するのが適切な防除法であろう。
補足:チークスケルトナイザー(Paliga machoeralis WalkerおよびP. damastesalis Walker)
前種のほかにチークで大発生する食葉性害虫としてはチークスケルトナイザー(teak skeletonizer)がある。インド、ミャンマー、タイ、インドネシアで古くから知られてきたが、最近チーク造林が盛んになったマレーシアでも食葉害が発生するようになった。しかし、大発生は雨季の後期に起こり、この時期のチークの材積成長はほぼ終了しているので、実質的には被害はないと結論されている。加害植物の記録はチーク類(Tectona spp.)以外ではシソ科のムラサキシキブ類(Callicarpa spp.)が知られているのみである。
チークスケルトナイザーは長らく “Eutectona machoeralis (Walker, 1895)”というメイガ科の1種として知られていたため、現在も多くの専門書でこの学名が使われている。ただし種小名は “machaeralis”と誤って綴られることが多い。また、インドネシアでは最近まで学名に “Pyrausta machoeralis (Walker, 1895)”が使用されることが多かった。しかし、属名EutectonaはPaligaのジュニアシノニムであるという指摘がなされている。また、現在の分類では従来のメイガ科をメイガ科(Pyralidae)とツトガ科(Crambidae)に分けており、Paliga属はツトガ科に所属する。したがって従来の多くの書籍・文献でメイガ科の“Eutectona machoeralis”とされてきた種は、ツトガ科のPaliga machoeralis (Walker, 1895)である。しかし、さらに同定上の問題があり、チークスケルトナイザーとされる種は少なくとも一部で誤同定されていることが最近判明し、インドネシアのジャワ、マレーシアの半島部とサバ州、タイでチークを加害している種は近縁種Paliga damastesalis (Walker, 1895)であることが確認された。他の地域でチークスケルトナイザーとされている種については今後正確な同定が必要である。Paliga machoeralisとP. damastesalisは似ており、ともに全体淡黄褐色で翅開長20〜25 mm。Paliga damastesalisでは前翅に赤色の波線状斑紋が比較的明瞭に現れ、後翅には斑紋がないのに対し、P. machoeralisでは前後翅とも灰色の波線状斑紋を持つ。生態についての過去の報告はどちらの種についてか明らかではないが、メスは1〜2週間生存して平均203〜374卵、最高500〜550卵を1個ずつ葉に産む;幼虫は緑色で成長すると約20 mm;葉脈を残して葉肉を食害する(英名の由来);葉を巻いた中や落葉下に繭を紡いで蛹化する。低緯度では休眠せず年間14世代までの発生が可能とされ、北インドでは幼虫で越冬休眠し、年間10世代の発生に留まると報告されている。
2.チビミノガ(Pteroma plagiophleps Hampson)


分類・分布
ミノガ科(Psychidae)の小型種で、幼虫はミノムシである。通常標記の学名が当てられているが、Pteroma属の分類は再検討を要するとの意見もある。インドからフィリピンまでの南アジア、東南アジアに分布する。
加害樹種
多食性で非常に多くの植物を食害する。造林樹種ではモルッカネム、アカシア類、ユーカリ類、チーク、キダチヨウラク等を食害し、マングローブ林や都市の庭園樹での発生も多い。
形態・生態
オス成虫は有翅の蛾で全体薄墨色、翅開長は15 mm程度、メスは無翅で生涯ミノの中にとどまる。メスは成虫になっても蛹の殻の中にいてミノに飛来したオスと交尾し、卵は自身の蛹殻内に産まれる。孵化した幼虫は外へ出て糸を吐いて風に乗り分散する。1メスあたり100〜200個体の幼虫が生まれる。成長途中の幼虫は円錐形のミノを作り、老熟幼虫のミノの長さ10〜15 mm、幼虫期間は約2ヶ月。老熟するとミノを紡錘形にして絹糸で懸垂して中で蛹化する。蛹期間は約2週間。1世代は約3ヶ月で通年幼虫が見られる。
被害
被害が多いのはモルッカネムで、理由は不明であるが他の樹種では大発生はあまり見られない。若齢幼虫は初め葉を食べているが、成長した幼虫は樹皮も剥ぎ取るように食べる。このため、大発生すると被害木の衰弱が著しく、枯れる木が続出する。しかし、被害が1〜数ヘクタールの狭い範囲に限って集中するため、連続した大面積の被害にはならない。これは孵化幼虫の分散様式とも関係しているのであろう。大発生は散発的で、季節性は不明瞭であるが乾季に多いという報告もある。孵化幼虫の風による分散以外に移動手段がない本種では、1ヘクタール程度の規模であっても大量の幼虫が突然出現することは考えにくいので、まず小規模な高密度パッチが発生し、1〜2世代かけて大発生に発達するのではないかと考えられるが、調べられていない。大発生が収束する過程では病気の流行や寄生蜂の寄与が大きいという報告がある。
防除
被害が小面積に留まるためか、本種による被害に対する関心は一般に低く、防除研究は実験室での化学薬剤の効果判定試験がわずかにあるだけで、ほとんど行われていない。幼虫がミノの中にいるので、化学的防除を行うのであれば接触剤よりも食毒剤・浸透剤の方が有効であろう。上述のようにおそらく大発生に先行する小面積の集中的な被害があると予想されるので、この段階で防除を行えば大発生を予防できると考えられるが、このような観点からの防除研究は行われていない。
補足:タイワンキチョウ(Eurema blanda Boisduval)シロチョウ科(Pieridae)
モルッカネムで大発生する食葉性害虫としては前種よりもタイワンキチョウが注目されることが多い。タイワンキチョウには同属近縁種が多く、特にキチョウ(ミナミキチョウ)Eurema hecabeとよく混同される。E. hecabeによるとされる食葉被害はほとんどが本種によるものである。中国南部、東南アジア、南アジア、ニューギニア、ビスマルク諸島、ソロモン諸島、オーストラリア北東部に広く分布する。成虫は前後翅とも黄色く外縁に黒帯があるが、オスでは後翅の黒帯が狭くなるか、ほとんど消失する。幼虫は黄緑色の体に黒い頭部を持つ。卵は展開していない葉芽に十数個から数十個の卵塊で産まれ、幼虫は集合して葉を食害する。蛹化もしばしば集合した状態で行われる。比較的高い位置(通常地上2 m以上)に産卵する。造林樹種ではモルッカネム(別名:センゴン、アルビジア、ファルカタ)(Falcataria moluccana (Miq.) Barneby & J.W.Grimes (=Paraserianthes falcataria (L.) Nielsen))、ネムノキ類(Albizia spp.)を加害し、ほかに、キンキジュ(Pithecellobium dulce)、ベニゴウカン類(Calliandra spp.)等のマメ科植物への加害が知られているが、Acacia mangiumやギンネム類(Leucaena spp.)は加害しない。
モルッカネム造林地で雨季の後半に大発生することがよくある。若いモルッカネムの林地で大発生が多いと述べている報告もあるが、むしろ比較的大径(胸高直径50cm以上)のモルッカネムが多い造林地では極端な大発生が多く、森林再生や水源涵養など産業目的以外で植林して伐採しないまま年数が経過した林地では毎年大発生が起こることも珍しくない。雨季に個体数が増えるのは産卵対象となる新葉の生産が降雨によって増加するためで、雨季を通じて増殖が続く結果雨季後半に大発生になる。大発生時には造林地全体の葉がなくなるなど食葉害が甚だしいことや発生する成虫数が夥しいことから、本種の大発生を見て被害を懸念する報告は多く、実際枝が枯れたり木全体が立ち枯れたりすることもあるが、そのような被害は稀である。成長阻害も(起きている可能性はあるが)問題視されたことはなく、慣行の短伐期で経営される生産林でも通常防除は行われていない。
参考文献
- Chey, V. B. (1996) Forest Pest in Sabah. Sabah Forest Department, Sandakan, 111 pp., 119 figs.
- Intachat, J. (1998) The identity of a Malaysian teak skeltoniser, Paliga damastesalis Walker (Lepidoptera: Pyralidae). Journal of Tropical Forest Science 10: 561-563.
- Irianto, R.S.B., K. Matsumoto and K. Mulyadi (1997) The yellow butterfly species of the genus Eurema Hubner causing severe defoliation in the forestry plantations of Paraserianthes falcataria (L.) Nielsen, in the western part of Indonesia. JIRCAS Journal 4: 41-49.
- Mathew, J. L. V. 1986. Variation in the wing venation of Pteroma plagiophleps Hampson (Lepidoptera: Psychidae). J. Res. Lepid. 24:359-363.
- 松本和馬 (2000) インドネシアのマメ科早生樹とその虫害問題.熱帯林業 47: 11-23.
- Mehlig, U. and M.P.M. de Menezes (2005) Mass defoliation of the mangrove tree Avicennia germinans by the moth Hyblaea puera (Lepidoptera: Hyblaeidae) in Equqtorial Brazil. Ecotropica 11: 87-88.
- Nair, K.S.S. (2007) Tropical forest insect pests: ecology, impact and management. Cambridge University Press, Cambridge.
- Nair, K.S.S., V.V. Sudheendrakumar, R.V. Varma and K.C. Chacko (1985) Studies on the seasonal incidence of defoliators and the effect of defoliation on volume increment of teak. Kerala Forest Research Institute Research Report 30: 1-78.
- Nair, K.S.S. and G. Mathew (1988) Biology and control of insect pests of first-growing hardwood species. Kerala Forest Research Institute Research Report 51: 1-8.
- Nair, K.S.S., Bijiu Babjan, T.V. Sajeev, V.V. Sudheendrakumar, M.I.M. Ali, R.V. Varma and K. Mohandas (1996) Field efficacy of nuclear polyhedrosis virus (NPV) for protection of teak against the defoliator, Hyblaea puera Cramer (Lepidoptera: Hyblaeidae). Journal of Biological Control 10: 79-85.
- Nair, K.S.S., R.V. Varma, V.V. Sudheendrakumar, K. Mohandas, M.I.M. Ali (1998) Management of the teak defoliator (Hyblaea puera) using nuclear polyhedrosis virus (NPV). Report of the research project KFRI 140/92 (Project BF/TF/05/02/91, sponsored by the Department of Biotechnology, Govt. of India) January 1992 to December 1995. Kerala Forest Research Institute Research Report 151: 1-64.
- Pres-Filho, O., A. Dorval, E. Berti-Filho (2002) Occurrence of teak defoliator Hyblaea puera (Cramer, 1777) (Lepidoptera: Hyblaeidae) in teak (Tectona grandis L.f.) in Brazil. Bragantia, Campinas 61: 59-60. (Portuguese with English abstact)